For more than a century, Neanderthal Man dragged through the collective imagination a reputation as a heavy meat-eater, a thick-set brute crouched around a fire, tearing apart with bare teeth the slab of mammoth he had just brought down. This image had the force of the obvious on its side: in the caves these hominins occupied, one found heaps of herbivore bones, tools made for cutting and deboning, and isotopic analyses that placed Neanderthal at the very top of the food chain, alongside the great carnivores. Yet over the past fifteen years, this portrait has cracked on every side. The tartar wedged between his molars, the fossilised excrement abandoned at the back of caves, and now the fine-grained analysis of his genome tellTellAn artificial mound formed by the accumulation of successive layers of settlement remains at the same spot, typical of the Near East. Each destruction-rebuilding event adds a stratum.→ a different story: that of an opportunistic omnivore whose plate was far more varied than anyone had believed, and which probably counted, among its resources, foods that most of us today regard with disgust or indifference, insects.
A recent paleogenetic study, published in the journal Science Advances under the title "Genetic dietary adaptation in Neandertal, DenisovanDenisovanAn extinct human population, cousin of the Neanderthals, identified in 2010 from the DNA of remains in Denisova Cave (Siberia).→ and Sapiens", shed unexpected light on this question. By comparing genes linked to metabolism and digestion in Neanderthals, Denisovans and Homo sapiensHomo sapiensThe present-day human species, which emerged in Africa around 300,000 years ago, the only surviving human lineage after the extinction of Neanderthals and Denisovans.→, the researchers brought to light genetic signatures suggesting that our vanished cousins were better equipped than we are to digest chitinChitinA rigid molecule forming the exoskeleton of insects; digesting it requires specific enzymes.→, the compound that forms the shell of insects. In other words, where the modern human body has partly turned its back on entomophagy, Neanderthal would have kept the digestive machinery for it. This article sets out to disentangle what science actually says, starting from the cliché of the carnivorous Neanderthal and arriving at a broader reflection on the dietary ecology of hominins and on what eating, in the PalaeolithicPalaeolithicThe oldest and longest period of prehistory (c. 3.3 Ma–12,000 BC), defined by chipped stone tools and a hunter-gatherer way of life.→, really meant.#s2

The cliché of the carnivorous Neanderthal
To understand where the idea of a meat-eating Neanderthal comes from, one must go back to the first great syntheses on his diet. From the end of the twentieth century, isotopic analyses of the nitrogen contained in the collagen of his bones yielded spectacular values. The ratio between heavy and light nitrogen isotopes rises with each link in the food chain: the higher one stands in the pyramid of predators, the higher this ratio. Neanderthal bones displayed signatures comparable to, or even higher than, those of the wolf, the hyena or the cave lion. The conclusion seemed self-evident: Neanderthal was a hypercarnivore, drawing most of his protein from the flesh of large herbivores such as reindeer, horse, bison or mammoth.#s3
This reading also rested on archaeology. Neanderthal sites overflow with game bones bearing cut marks, traces of fracturing to extract marrow, signs of cooking. The technical toolkit, scrapers, points, denticulates knapped according to the Levallois method, seemed tailor-made for butchery. Anatomy itself appeared to corroborate this specialisation: a stocky, muscular, calorie-hungry body, a flared rib cage that could house an oversized liver and kidneys, presumed to handle the excess nitrogen typical of a very meaty diet. Everything pointed to making Neanderthal the great predator of Europe's glacial steppes.
But this reasoning suffered from a major bias. Nitrogen isotopes measure the share of animal protein in the diet, and they saturate quickly: beyond a certain threshold, one can no longer distinguish a moderate meat-eater from an absolute hypercarnivore. Above all, they say nothing about foods that are poor in nitrogen or consumed in small quantities, plants, fungi, and precisely those small prey that insects are. A whole portion of the diet remained invisible to the instrument. Moreover, the isotopic signal mainly reflects the origin of the protein, not overall caloric intake; yet foods such as tubers, seeds or larvae can supply a great deal of energy without weighing heavily in the nitrogen balance. The carnivorous Neanderthal was, in part, an artefact of method.
Added to this was a cultural framing. The image of the heroic hunter fitted well with the idea one had of survival in a glacial setting, and it valued meat as the engine of brain evolution. This "meat hypothesis" long dominated paleoanthropology, relegating gathering and so-called minor resources to the rank of negligible supplement. Yet present-day hunter-gatherersHunter-gatherersA way of life based on hunting, fishing and gathering wild resources, without farming or herding; it dominated almost the whole of human history.→ teach us the opposite: in many societies, it is plant resources and small prey, collected daily, that ensure food security, while big-game hunting, more uncertain, provides irregular returns. Nothing prevented applying this pattern to Neanderthal.
What dental tartar and coprolites reveal
It is worth recalling how this image spread far beyond the circle of specialists. Over the course of the twentieth century, schoolbooks, museums and popular culture froze Neanderthal in the posture of the mammoth hunter, spear in hand, draped in furs. This iconography was not neutral: it served a narrative in which mastery of big-game hunting marked the accession to humanity, and in which meat, a dense source of protein and fat, appeared as the fuel of cerebral expansion. In this scheme, everything that fell under patient gathering, the scraping of roots or the collecting of larvae was pushed into the shadows, judged too trivial to deserve attention. The dietary cliché thus took part in an implicit hierarchy of behaviours, where predation rhymed with nobility and gathering with banality.
Yet real subsistence economies cannot be reduced to this opposition. Among the hunter-gatherers documented by ethnography, caloric intake often comes in large part from modest but reliable resources, collected by women, children and the elderly, while big-game hunting, prestigious and uncertain, covers only a fraction of daily needs. Transposed to the Palaeolithic, this observation cautions against confusing what the remains make visible, the large, well-preserved bones, with what really made up the daily diet. Insects, plants and small prey, unlikely to leave durable traces, probably weighed far more heavily in the balance than the bone heaps of the caves suggest.
The turning point came from sources that had long been neglected: dental tartar and coprolites. Tartar, that mineralised plaque which accumulates on teeth for lack of oral hygiene, is a genuine time capsule. As it calcifies, it traps plant microfossils, starch grains, phytoliths, those tiny silica concretions that plants manufacture, as well as fragments of tissue, spores, fibres and organic molecules. Under the microscope and thanks to analytical chemistry, this deposit yields a snapshot of what passed through the individual's mouth, sometimes tens of thousands of years earlier.#s3
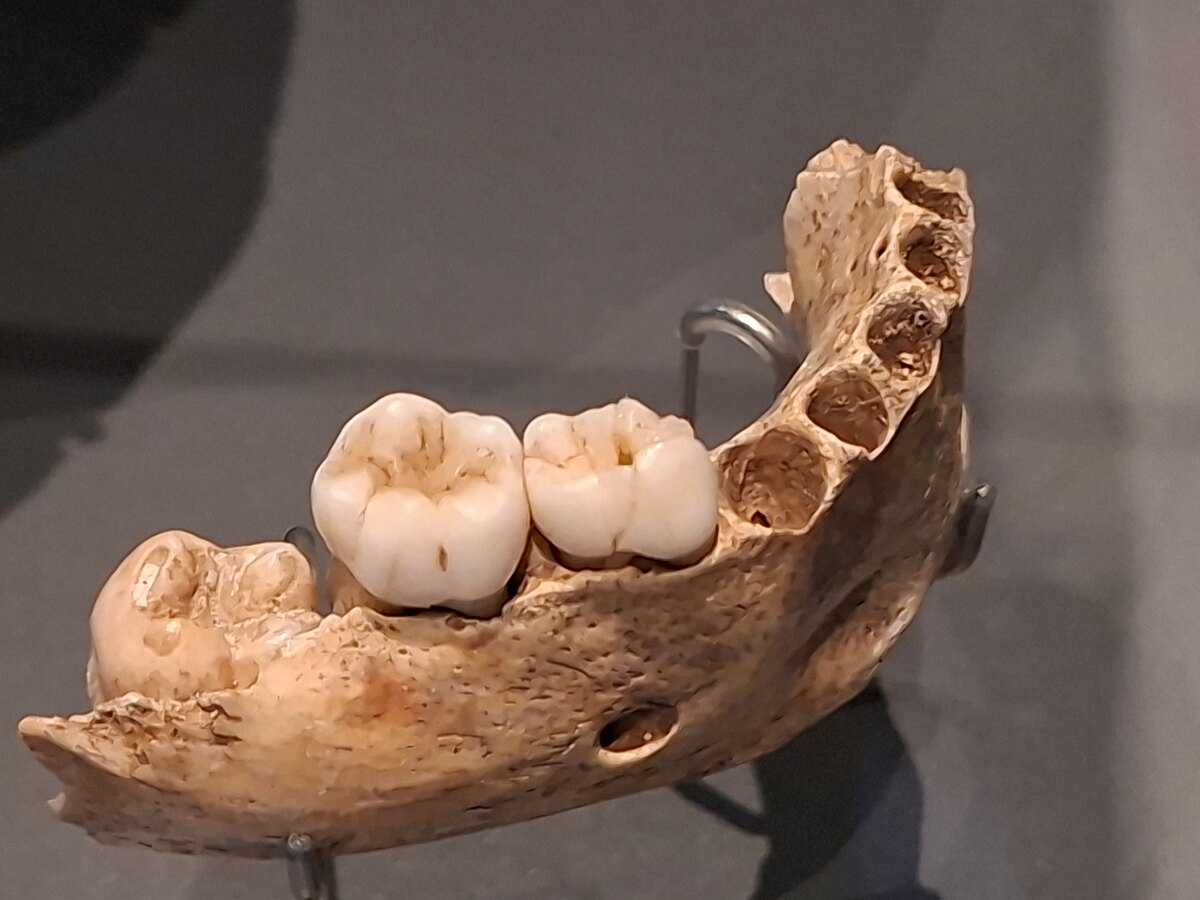
Analyses of Neanderthal tartar held surprises. At several sites, cooked starch grains were identified, coming from grasses, legumes, even tubers, evidence of regular consumption of plants sometimes prepared over fire. At El Sidrón, in Spain, the tartar of Neanderthal individuals even yielded compounds associated with bitter plants of no obvious nutritional value, which some researchers interpreted as a medicinal use or, more cautiously, as the trace of plants consumed for other reasons. In other contexts, residues of fungi, pine nuts and mosses were found. The menu was proving far more plant-based and far more varied than the isotopes had suggested.
Coprolites, those fossilised excrements, are another precious window. When well preserved, they contain biomarkers characteristic of what has been digested. At the site of El Salt, in Spain, the analysis of sediments interpreted as Neanderthal faecal matter revealed the simultaneous presence of coprostanol, a marker of the digestion of animal cholesterol, and of plant-derived sterols, a sign of a mixed diet combining meat and plants. These results, obtained on residues about fifty thousand years old, reinforced the idea of an effective omnivory rather than a strict carnivory.
One category of food remained particularly hard to detect: insects. Their chitin shell, their soft tissues, their small hard parts preserve poorly and often go unnoticed in classic analyses. Yet fragments of exoskeletons, beetle elytra or larval remains do sometimes appear in the coprolites of more ancient hominins and in other primates. The absence of massive direct evidence is not evidence of absence: it first reflects a bias of preservation and detection. It is precisely this documentary void that the genetic approach came to fill, by seeking not the insects themselves, but the trace of the adaptation that allowed them to be digested.
The genetic study: chitinases and chitin digestion
Chitin is one of the most abundant polymers on the planet. It is what stiffens the shell of insects and crustaceans, the wall of many fungi, certain structures of worms and molluscs. To exploit a whole insect as food, one must be able, at least partly, to overcome this envelope. This is the role of chitinases, enzymes that cleave the long chains of chitin into assimilable fragments. In mammals, the main one is acid chitinase, or AMCase, encoded by the CHIA gene. It is found notably in the stomach and the respiratory tract, where it takes part in the breakdown of ingested chitin and in certain immune responses.#s2
The study published in Science Advances compared the versions of these metabolic genes in Neanderthals, in the Denisovan and in Homo sapiens, drawing on the high-quality ancient genomes now available. The guiding idea was simple: if a group preserves a complete enzymatic apparatus for digesting chitin, it is probably because this food held a real place in its diet; conversely, the erosion or inactivation of these genes signals a turning away from this resource. The researchers therefore scrutinised the state of the CHIA gene and other loci associated with digestion, assessing whether they were functional, partly degraded or silent.#s1
The result is striking. In Neanderthals and in the Denisovan, the genetic apparatus linked to chitin digestion appears better preserved, more complete, than in the average modern Homo sapiens. This suggests that these hominins had a more robust enzymatic capacity to process insects, and therefore that entomophagy was very likely part of their ordinary dietary ecology. The signature is not limited to chitinase: other metabolic genes, involved in the processing of lipids or various nutrients, show variants consistent with a broad diet, exploiting diversified animal sources, from small prey to large herbivores.
One must stress the methodological elegance of this approach. Rather than waiting for the improbable discovery of a coprolite stuffed with elytra or a tooth studded with insect fragments, it interrogates the genome as an archive of dietary behaviour. The logic is that of evolutionary genetics: a gene remains functional over the long term only if it confers an advantage, that is, only if it is used. The conservation of an active chitinase arsenal in Neanderthals and Denisovans thus becomes an indirect but powerful clue to regular insect consumption. Paleogenetics is no longer content to reconstruct lineages or episodes of hybridisationHybridisationCrossing between two distinct species or lineages, such as Homo sapiens and Neanderthals, leaving a trace in the genome.→ between populations; it now illuminates entire swathes of the way of life.
One last element deserves attention: cooking. The mastery of fire, attested in Neanderthals, radically changes the yield of foods. Heating a tough plant, a bitter root or an insect makes its nutrients more accessible, neutralises certain toxins and eases digestion. The presence of cooked starches in Neanderthal tartar confirms this. Cooking also lowers the digestive cost of chitin and insect tissues: grilled, locusts or larvae become more digestible and more palatable. The enzymatic capacity inscribed in the genes therefore did not work alone; it combined with a culinary know-how that multiplied the range of exploitable resources. Tartar, coprolites and hearths together sketch a Palaeolithic cuisine far more elaborate than once imagined.

A faculty lost in Homo sapiens
If Neanderthals and Denisovans were well equipped to digest chitin, what about us? In contemporary Homo sapiens, the picture is more contrasted. The CHIA gene remains present and expressed, but its activity varies greatly from one individual and population to another, and several human lineages carry variants that reduce its efficiency. Overall, the average enzymatic capacity to break down chitin seems to have receded in modern humans compared with what the Neanderthal and Denisovan genomes suggest. Part of the digestive toolkit inherited from our common ancestors appears, in Sapiens, to be loosening.
How can this retreat be explained? The most natural hypothesis links the state of a gene to the use made of it. When a resource ceases to be central to a population's diet over long periods, the selective pressure that maintained the associated genes relaxes, and these genes can accumulate mutations without immediate consequence, until they lose part of their function. If entomophagy became marginal in some Sapiens populations, in favour of other protein sources, fishing, herding, agricultureAgricultureThe cultivation of plants and production of food by working the soil, which emerged in the Neolithic in the Near East and independently elsewhere; it radically transformed human societies.→, maintaining an enzymatic apparatus optimised for chitin no longer brought a decisive advantage. The faculty did not vanish all at once; it eroded, unevenly, according to regional diets.
One must beware of any hasty generalisation. "Homo sapiens" is not a homogeneous entity: hundreds of millions of people, even today, consume insects regularly and apparently without any notable digestive difficulty. The variability of the CHIA gene from one population to another reflects precisely this diversity of diets. What the genetic comparison shows is an average tendency, a statistical contrast between very well-equipped ancient human lineages and a Sapiens whose equipment, on average, has partly weakened. It is less the loss of a capacity than its fragmentation and its uneven conservation across the species.
This observation fits into a broader phenomenon: that of the genes of taste, digestion and metabolism that bear the imprint of our ancestors' dietary choices. Lactose tolerance in adulthood, adaptation to starch-rich diets through the multiplication of amylase genes, variations in the perception of bitterness: so many examples where diet has sculpted the human genome in a few thousand generations. Chitinase joins this gallery. It reminds us that our genetic heritage is not fixed but records, gene after gene, the history of what we have eaten.
One may extend the reflection by asking what "digesting an insect" concretely means. Eating a grasshopper or a larva is not only about absorbing the animal's soft flesh, rich in protein and fat; it is also about ingesting its chitin exoskeleton. Part of this chitin passes through the digestive tract without being assimilated and plays the role of fibre, but a fraction can be broken down by chitinases and contribute to nutrition, or modulate the gut flora and immune response. An efficient enzymatic apparatus therefore does not merely make the insect edible; it optimises its nutritional yield and softens any digestive discomfort. For a population whose insects were a regular resource, having effective chitinases represented a tangible advantage.
This reasoning also illuminates the notion of a conserved capacity. Keeping a gene fully functional has a cost: it must be maintained, expressed, protected from deleterious mutations by selection. If this maintenance persists over tens of thousands of years, it is because the advantage it provides offsets that cost. The good preservation of chitinases in Neanderthals and Denisovans is therefore not a neutral accident, but the sign of a function actively sustained by use. Conversely, the relaxation observed on average in Sapiens reflects a dietary history in which, for many populations, this function ceased to be critical. The genome thus records, in negative, the gradual shift of eating habits.
It is also worth dwelling on the sheer scale and predictability of the insect resource in a glacial Europe. Far from being a marginal scattering of bugs, certain insect populations form dense, seasonal, locatable concentrations: swarms of locusts, mass emergences of winged termites or aquatic larvae, beehives and wasp nests, wood-boring grubs hidden under bark. For a homininHomininMember of the subtribe Hominina, comprising the human lineage (Homo, Australopithecus, Paranthropus…) but excluding orangutans and gibbons. The term progressively replaces "hominid" in its narrow sense.→ who knew the landscape intimately, these were appointments to be kept, predictable harvests as reliable as a fruiting tree or a salmon run. Exploiting them required no weapons and little risk, only knowledge and timing. In an economy where every calorie counted and where the failure of a hunt could mean days of hunger, such a steady, low-cost supplement would have been precious, all the more so when grilled over the very fire whose mastery the Neanderthals had long since acquired.
Entomophagy in human evolution
Placed in the long term, insect consumption by hominins is nothing exotic: it is rather the opposite that calls for explanation. Among primates, entomophagy is extremely widespread. Many monkeys and prosimians supplement their diet with insects, and some, like tarsiers or various galagos, even make them their main resource. Chimpanzees, our closest living relatives, fish for termites with twigs, dig out ants, crunch larvae and caterpillars. This practice provides high-quality protein, lipids, vitamins and minerals in a form that is easy to collect, often abundant and predictable with the seasons. The insect is, for a primate, a first-rate food resource.
It would therefore be surprising if the human lineage had been an exception. Everything indicates, on the contrary, that our distant ancestors, australopithecines and the first representatives of the genus Homo, continued to exploit this windfall. Analyses of dental microwear and certain isotopic studies on ancient hominins from southern AfricaAfricaThe cradle of humankind: the continent where the first hominins appeared, then Homo sapiens around 300,000 years ago, before the expansion to the rest of the world.→ have suggested a notable share of social insects, such as termites, in their diet. The ability to locate, extract and consume insects was not a late innovation but a continuous inheritance, passed down from generation to generation since long before the appearance of Neanderthals.
Present-day human populations offer an illuminating mirror. Entomophagy is practised on every inhabited continent, by extremely diverse societies. In sub-Saharan Africa, mopane caterpillars, winged termites and locusts are appreciated and traded foods. In Southeast Asia, grasshoppers, silkworms, water bugs and various larvae fill the stalls. In Latin America, ants, grasshoppers and agave larvae feature in traditional dishes that are sometimes highly prized. In Australia, Aboriginal peoples have for millennia consumed the larvae known as witchetty grubs. Far from being a curiosity, eating insects is one of the oldest and most universal human ways of feeding oneself.
What modern Western repugnance tends to obliterate is the ecological and nutritional efficiency of this resource. Insects display an excellent yield: it takes far fewer resources to produce a gram of insect protein than a gram of beef protein. They are rich in essential amino acids, in iron, in zinc, in good fatty acids. For a Palaeolithic hunter-gatherer, a swarm of locusts, a nest of larvae or a colony of termites represented a concentration of energy that could be mobilised quickly, without the risk or the energy expenditure of a big hunt. That Neanderthal knew how to exploit this opportunity seems, in this light, perfectly logical.
Ecological implications: a varied and flexible diet
The main lesson of this work is not that Neanderthal ate insects, a fact, after all, expected of an omnivorous primate, but that the representation of his dietary ecology must be entirely revised. The portrait of the single-minded predator gives way to that of a flexible opportunist, able to adjust his diet to the available resources according to place, season and circumstance. This flexibility is precisely what characterises robust species, capable of colonising varied environments and of weathering periods of climatic instability.
Comparing the peoples who still practise entomophagy also reveals the cultural dimension of this diet. Where some societies have integrated insects into their daily cuisine, developing techniques of harvesting, preservation and preparation, others have gradually set them aside, to the point of making them a dietary taboo. The revulsion that insects inspire in much of the Western world is in no way natural or universal: it is a recent cultural construction, linked to the rise of livestock farming, urbanisation and a certain representation of cleanliness. Understanding this helps us reread the past without projecting our disgusts: for Neanderthal as for most humans in history, crunching an insect was nothing repugnant, it was a nutritional obviousness.
Now Neanderthals occupied an immense range, from the Iberian Peninsula to the edges of Siberia, by way of the Near EastNear EastA region of western Asia (Levant, Mesopotamia, Anatolia, Iran), cradle of the Neolithic revolution, agriculture, the first cities and writing.→, over several hundred thousand years marked by abrupt glacial oscillations. Such longevity and such geographic extension are hardly compatible with a narrow and rigid diet. On the contrary, they presuppose a broad dietary palette: large herbivores when hunting allowed, but also small game, birds, fish and shellfish on the coasts, plants, fungi, dried fruits, and insects as opportunities arose. The diversity of the menu is the logical counterpart of the diversity of the environments crossed.
This revision has consequences for the way we conceive of Neanderthal's ecological intelligence. Exploiting such a varied range of resources requires knowledge: knowing the reproductive cycles of insects, spotting colonies, knowing where and when to gather, how to prepare plants that are sometimes toxic in the raw state. This presupposes a memory of places, a transmission of techniques, a capacity for planning. Far from the crude brute of the cliché, there emerges the image of a subtle actor in his environment, endowed with a detailed knowledge of the biology surrounding him. Omnivory is not only a matter of digestion; it is also a matter of cognition.
This dietary flexibility joins other recent findings about Neanderthal: use of fire, manufacture of adhesives, care for the injured, perhaps symbolic practices. The overall picture is that of a fully-fledged humanity, endowed with adaptive capacities comparable to our own. The varied diet, once underestimated, becomes one of the arguments of this rehabilitation. Eating everything, and knowing how to do so, is in itself a form of intelligence about the world.
Link with the disappearance of Neanderthal, or not
As soon as one mentions Neanderthal's diet, the question of his disappearance is never far. A long-popular hypothesis held that his supposed carnivorous specialisation made him vulnerable: too dependent on big game, he would have been unable to adapt to climatic upheavals and to competition from the more versatile Sapiens. If this specialisation turns out to be largely illusory, then a whole part of this scenario collapses. An omnivorous and flexible Neanderthal was not trapped by a narrow diet.
One must therefore be cautious. The fact that Neanderthal had a broad dietary repertoire, insects included, argues against the idea of an extinction through ecological rigidity. His disappearance, some forty thousand years ago, more likely results from a bundle of causes: fragile demography and scattered populations, gradual interbreeding with Sapiens, of which our genomes still bear the trace, dynamics of competition and absorption rather than brutal replacement. Diet does not appear as the Achilles' heel that some have wished to see in it.
One could even reverse the argument. If Neanderthal retained a superior enzymatic capacity to digest chitin, it is perhaps because he exploited this resource with an efficiency at least equal to that of Sapiens. This does not make entomophagy a decisive survival factor, but it ruins the idea of a Neanderthal dietary inferiority. The two lineages shared the same omnivorous base inherited from their common ancestors; on the ground of diet, nothing indicates that one was structurally better armed than the other.
The lesson is that one must resist the temptation to make every newly discovered trait the cause of Neanderthal's disappearance. Paleoanthropology has multiplied these single-cause explanations, climateClimateThe long-term average atmospheric conditions of a region; its variations (glaciations, aridifications) shaped migrations, agriculture and the collapse of prehistoric societies.→, brain, language, diet, often later refuted. Neanderthal's varied diet enriches our understanding of his way of life without thereby providing the key to his end. It invites us instead to separate these two questions: how he lived, and why he disappeared, belong to distinct logics.
It is worth measuring, too, how far this reappraisal carries us from the old evolutionary narratives. For decades, the story of human origins was told as a triumph of the hunter: the more meat, the bigger the brain, the more human the creature. Insects, plants and patient gathering had no place of honour in that tale. Reinstating them does not diminish Neanderthal; it makes him more recognisable. The being who reads a landscape for its hidden larvae, who times the emergence of termites, who grills a handful of grasshoppers alongside a cut of horse, is not a failed experiment on the road to us, he is a competent forager solving the same daily problem every human society has faced. The varied diet is, in this sense, less a curiosity than a sign of full membership in the human story.
Methodological limits
Like any advance, these results call for critical examination. The first limit lies in the indirect nature of the genetic evidence. To observe that a chitinase gene is well preserved in Neanderthals suggests insect consumption, but does not prove it in the way an elytron fragment in a dated coprolite would. The gene attests a capacity, not necessarily an effective behaviour nor its frequency. Other evolutionary pressures, immune responses, interactions with pathogens, a role in the respiratory tract, can contribute to the maintenance of these genes independently of diet. The inference is solid, but it remains an inference.
A second difficulty: the quality and representativeness of ancient genomes. A few sequenced individuals do not sum up the diversity of populations spread over hundreds of thousands of years and thousands of kilometres. Ancient DNAAncient DNAFragments of DNA preserved in old remains (bones, sediment); their sequencing identifies species and traces vanished lineages.→ is fragmented, contaminated, sometimes incomplete; reconstructing the precise functional state of a gene from such material demands precautions and carries a margin of uncertainty. Conclusions drawn from a small sample must be regarded as tendencies to be confirmed, not as definitive truths valid for all Neanderthals.
A third point: biases of preservation and detection. The absence of insect remains at most Neanderthal sites does not mean that none were consumed, but it also deprives the genetic hypothesis of a direct and abundant confirmation. The methods capable of spotting chitin biomarkers or insect microremains in tartar and sediments are still under development. As long as this material evidence remains rare, the debate will retain a share of reasoned speculation, in which genetics offers a strong lead that field archaeology has not yet fully corroborated.
The flexibility of the diet finally sheds light on the question of resilience in the face of crises. When an environment deteriorates, abrupt cooling, scarcity of game, retreat of forests, a species with a narrow diet sees its resources collapse all at once, whereas a generalist species can fall back on backup solutions. Insects and small prey typically belong to this fallback repertoire: abundant, renewable, accessible without much equipment, they offer a safety net when big fauna becomes scarce. That Neanderthal retained the ability to exploit them suggests that he too had these margins of manoeuvre in the face of climatic instability. The diversity of the diet is not merely a gastronomic curiosity: it is a survival asset.
Finally, one must be wary of overly seductive narratives. The image of an insect-crunching Neanderthal holds strong media appeal, and the temptation is great to over-interpret subtle genetic signals. Rigour requires distinguishing what is established, omnivory, the diversity of the diet, the differential conservation of certain genes, from what remains hypothetical, the exact quantitative importance of insects in the diet, its regional variations, its modes of preparation. It is at this price that scientific progress does not degrade into mere storytelling.
Prospects
The coming years should considerably refine this picture. On the genomic side, the increasing number of high-quality Neanderthal and Denisovan genomes will allow a finer mapping of the variability of metabolic genes, an assessment of whether the good conservation of chitinases is general or specific to certain populations, and a linking of these signatures to precise ecological contexts. Functional genomics, able to test in the laboratory the real activity of the enzymes encoded by these ancient genes, will provide a direct measure of digestive capacity, beyond the mere presence of the gene.
On the archaeological side, the refinement of analyses of dental tartar, coprolites and sedimentary residues promises to flush out traces hitherto undetectable. Lipid biomarkers specific to chitin, the search for insect DNA in deposits, very-high-resolution microscopy could one day provide the direct material evidence that genetics leads us to anticipate. The systematic crossing of these approaches, genetic, isotopic, microfossil, biomolecular, is the most promising path to reconstruct, resource by resource, the real menu of the Neanderthals.
It is also worth situating these discoveries within the long march of paleogenetics, a discipline that has overturned our vision of hominins in less than two decades. The sequencingSequencingReading the order of the bases (A, T, G, C) of a DNA molecule; high-throughput sequencing reads millions of fragments in parallel.→ of the Neanderthal genome, then the identification of the Denisovan from a single bone fragment, proved that one could read the past in molecules. Reconstructing a diet from the state of a few genes extends this ambition: making ancient DNA speak no longer only of lineages, but of behaviours, ecology, ways of life. Chitinase thus joins a bundle of clues, genes of immunity, of pigmentation, of metabolism, that, patiently assembled, give flesh back to vanished populations.
Beyond Neanderthal, this research outlines a broader programme: to reconstruct the dietary ecology of all hominins from the convergence of genetic and material clues. Understanding how each species exploited its environment, which genes accompanied these choices, how diet in turn shaped anatomy and physiology, this is a project that is only beginning. Chitinase is but one thread; pulling on it unwinds a whole history of the coevolution between humans and their food.
These prospects also have a contemporary resonance. As entomophagy is promoted as a sustainable answer to the challenges of world food supply, rediscovering its deep antiquity in the human lineage offers a useful light. Eating insects is not a futuristic novelty but a return to the source, a practice inscribed in our evolutionary history and in our genes. Paleogenetics, by exploring the most distant past, thus joins the most current debates on what we put on our plates.
Conclusion
The carnivorous Neanderthal, brute of the glacial steppes obsessed with meat, now belongs to the past of science. In his place stands the image of a subtle omnivore, exploiting a range of resources in which large herbivores rubbed shoulders with plants, fungi, small prey and, very probably, insects. The tartar of his teeth, the excrement he left and, more recently, the analysis of his genome converge towards this conclusion. The study of chitinases, by revealing in Neanderthals and Denisovans an apparatus better preserved than in Sapiens for digesting chitin, adds an elegant piece to this file: our vanished cousins were, on this point, better equipped than we are.
This rediscovery goes beyond a mere dietary anecdote. It forces us to think of Neanderthal as a sophisticated, flexible and knowledgeable ecological actor, whose success rests precisely on this capacity for adaptation. It also shifts the gaze upon ourselves: the partly lost faculty of easily digesting chitin testifies to the way our diets have, slowly, reshaped our genetic heritage. Between the insect crunched at the back of a cave fifty thousand years ago and today's debates on sustainable food runs a thin but real thread, that of an evolutionary history in which what we eat shapes what we are.
There remains, finally, an invitation to humility. Each generation of researchers believed it held, with its carnivorous, herbivorous, cannibal or cook Neanderthal, the definitive portrait of this hominin. The lesson of recent decades is that this portrait is constantly being nuanced, as new methods open new windows. Neanderthal entomophagy is not the last word; it is one more piece in an ever-unfinished mosaic. But it is a piece that, in its own way, brings us closer to our cousins: imagining them crouched near a fire, sharing grilled locusts as much as a haunch of reindeer, we recognise in them fully adapted, ingenious humans who are, at heart, astonishingly close to us.
L'entomologie archéologique est une discipline que peu de gens connaissent et cet article est une belle introduction. Les insectes se conservent dans des contextes archéologiques particuliers et leur étude renseigne autant sur l'environnement que sur les comportements humains. C'est un bon exemple pour montrer à mes élèves l'interdisciplinarité des sciences archéologiques.
La découverte de restes d'insectes associés aux sépultures néandertaliennes est une piste de recherche relativement récente mais très prometteuse. Les coléoptères en particulier sont d'excellents indicateurs environnementaux et taphonomiques. S'ils étaient consommés comme complément alimentaire ou avaient une valeur symbolique, cela enrichirait encore notre image de la complexité comportementale néandertalienne.