Il y a quelque chose de vertigineux dans l'idée que les milliards d'êtres humains répartis aujourd'hui sur tous les continents, des Inuits du Grand Nord aux Aborigènes d'Australie, des Andins aux Sibériens, descendent, pour l'essentiel de leur héritage génétique, d'une poignée de populations qui quittèrent l'AfriqueAfriqueContinent berceau de l'humanité : les premiers hominines y apparaissent, puis Homo sapiens il y a environ 300 000 ans, avant l'expansion vers le reste du monde.→ il y a environ soixante mille ans. C'est là le cœur de ce que la science nomme la « sortie d'Afrique », ou Out of Africa : le scénario, aujourd'hui solidement étayé, selon lequel notre espèce, Homo sapiensHomo sapiensEspèce humaine actuelle, apparue en Afrique il y a environ 300 000 ans, seule lignée humaine survivante après l'extinction de Néandertal et des Dénisoviens.→, est née sur le sol africain avant d'essaimer à travers le monde et de remplacer, partout, les autres humanités qui l'avaient précédée hors du continent.
Ce récit, qui paraît aujourd'hui presque évident, a longtemps été disputé. Pendant la seconde moitié du XXe siècle, deux écoles se sont affrontées : les partisans d'une origine multirégionale, qui imaginaient une évolution parallèle d'Homo sapiens sur plusieurs continents à partir de populations d'Homo erectus locales, et ceux d'une origine africaine récente et unique. C'est cette seconde hypothèse qui l'a emporté, portée d'abord par la paléogénétiquePaléogénétiqueÉtude de l'ADN ancien extrait de restes (os, dents, sédiments, parois) pour reconstituer le passé des populations.→, l'étude de l'ADN ancienADN ancienFragments d'ADN conservés dans des restes anciens (os, sédiments) ; leur séquençageSéquençageLecture de l'ordre des bases (A, T, G, C) d'une molécule d'ADN ; le séquençage à haut débit lit des millions de fragments en parallèle.→ permet d'identifier des espèces et de retracer des lignées disparues.→ et des génomes actuels, puis confirmée par la paléoanthropologiePaléoanthropologieScience qui étudie l'évolution humaine à partir des restes fossiles d'hominidés (os, dents, empreintes) et de leur contexte, pour reconstituer nos origines biologiques.→ et l'archéologie. Cet article retrace cette grande aventure : l'origine africaine de notre espèce, la chronologie des sorties successives, les routes empruntées vers le Levant et l'Arabie, la preuve génétique de l'Ève mitochondriale, l'énigme du goulot d'étranglement et de l'éruption du Toba, les métissages avec les Néandertaliens et les Dénisoviens, et enfin le peuplement effectif de la planète entière.
L'origine africaine d'Homo sapiens
Avant de pouvoir sortir d'Afrique, Homo sapiens devait d'abord y naître. Or, sur ce point précis, les vingt dernières années ont profondément renouvelé notre compréhension. Longtemps, on a cru que notre espèce était apparue en Afrique de l'Est, dans la région de la vallée du Grand Rift, il y a environ deux cent mille ans. Les fossiles d'Omo Kibish, en Éthiopie, datés d'environ cent quatre-vingt-quinze mille ans, et ceux de Herto, également éthiopiens, vieux de cent soixante mille ans, semblaient confirmer ce berceau oriental. Ces crânes, désignés sous les noms d'Homo sapiens idaltu pour Herto, présentent déjà l'essentiel des traits anatomiques modernes : un crâne haut et arrondi, un front vertical, une face réduite, un menton osseux.
Mais en 2017, la découverte révisée du site de Jebel Irhoud, au Maroc, a fait reculer cette date et déplacé la géographie du problème. Les restes humains de Jebel Irhoud, longtemps mal datés, ont été attribués à environ trois cent mille ans grâce à la datation par thermoluminescence des silex brûlés associés. Ces fossiles marocains, qui combinent une face déjà moderne avec un crâne encore allongé et archaïqueArchaïqueSe dit d'une population ou d'une forme humaine ancienne et aujourd'hui disparue (Néandertal, Denisova, lignées fantômes), par opposition aux humains anatomiquement modernes.→, ne représentent sans doute pas nos ancêtres directs au sens strict, mais ils témoignent qu'à l'aube de notre lignée, des populations à la morphologie proche de la nôtre vivaient déjà à l'extrémité nord-ouest de l'Afrique, à des milliers de kilomètres du Rift.
De cette accumulation de données est née une idée nouvelle, défendue notamment par la paléoanthropologue Eleanor Scerri : celle d'une origine « panafricaine » d'Homo sapiens. Plutôt qu'un berceau unique et localisé, notre espèce serait issue d'une mosaïque de populations dispersées à travers tout le continent, du Maroc à l'Afrique australe, parfois isolées par les déserts ou les forêts, parfois reconnectées au gré des oscillations climatiques. Ce serait de ces échanges intermittents, de ce brassage à l'échelle continentale, qu'aurait émergé peu à peu la morphologie et le génome de l'humanité moderne. Notre espèce ne serait pas tant le produit d'un lieu que celui d'un réseau, d'une métapopulation africaine.
Une ou plusieurs sorties ? La sortie précoce et la grande dispersion
La question des dates de sortie est l'une des plus débattues de la discipline, et elle s'est considérablement compliquée à mesure que de nouveaux fossiles affleuraient. Le schéma classique, encore largement valide pour expliquer l'ascendance des populations actuelles, place la grande dispersion entre soixante et cinquante mille ans. C'est cette vague-là, et elle seule, qui aurait laissé une empreinte génétique durable chez l'ensemble des humains vivant aujourd'hui hors d'Afrique. Mais les indices d'incursions plus anciennes se sont multipliés.
À Misliya, dans le mont Carmel en Israël, un fragment de maxillaire daté entre cent soixante-dix-sept et cent quatre-vingt-quatorze mille ans atteste qu'Homo sapiens avait atteint le Levant bien avant la grande sortie. Les grottes voisines de Skhul et de Qafzeh ont livré des sépultures de sapiens vieilles de cent à cent vingt mille ans. Plus loin encore, des dents découvertes à Daoxian, dans le sud de la Chine, suggèrent une présence d'humains modernes en Asie de l'Est il y a quatre-vingt mille à cent vingt mille ans. Enfin, en Arabie, le site de Al Wusta a livré une phalange humaine moderne datée d'environ quatre-vingt-cinq mille ans.
Comment réconcilier ces traces précoces avec l'ascendance presque exclusivement « récente » des populations actuelles ? La réponse la plus admise distingue deux types d'événements. D'une part, des sorties précoces, dès cent à cent vingt mille ans, qui auraient porté de petits groupes de sapiens jusqu'au Levant et peut-être jusqu'en Asie, mais qui se seraient pour l'essentiel éteintes ou résorbées, ne laissant qu'une contribution génétique marginale, voire nulle, chez les vivants. D'autre part, la grande dispersion d'il y a soixante mille ans, démographiquement victorieuse, dont descendent tous les non-Africains actuels. Les sorties précoces seraient ainsi des bourgeons avortés, les « répétitions générales » d'un exode dont seule la dernière fut couronnée de succès.
Les routes : le Levant et la « route sud » arabique

Pour quitter l'Afrique, deux grandes portes s'offraient à Homo sapiens. La première, la plus septentrionale, est le corridor levantin : en remontant la vallée du Nil puis en franchissant l'isthme de Suez, on débouche sur le couloir syro-palestinien, qui relie naturellement l'Afrique à l'Eurasie. C'est par là, sans doute, que sont passés les sapiens de Misliya, de Skhul et de Qafzeh. Cette route avait toutefois un inconvénient majeur : elle dépendait étroitement du climatClimatConditions atmosphériques moyennes d'une région sur le long terme ; ses variations (glaciations, aridifications) ont orienté migrations, agriculture et effondrements de sociétés préhistoriques.→. Aux périodes humides, le Sahara et le Levant se couvraient de savanes et de lacs, on parle de « Sahara vertSahara vertNom donné au Sahara durant la « période humide africaine » (env. 14 500 à 5 000 ans avant le présent), lorsque des pluies de mousson accrues y entretenaient lacs, rivières et savanes, rendant la région habitable avant son assèchement progressif.→ », et le passage devenait praticable ; aux périodes arides, le désert se refermait comme une barrière infranchissable.
La seconde porte, méridionale, est la fameuse « route du sud ». Elle suppose une traversée du détroit de Bab-el-Mandeb, à l'extrémité de la mer Rouge, là où la corne de l'Afrique n'est séparée de l'Arabie que par une trentaine de kilomètres. Lors des maxima glaciaires, l'abaissement du niveau marin, qui pouvait atteindre cent vingt mètres, réduisait encore cette distance et faisait émerger des îles et des hauts-fonds, rendant la traversée envisageable même pour des groupes ne disposant que d'embarcations rudimentaires. Une fois en Arabie, les migrants auraient longé les côtes de l'océan Indien, exploitant les ressources littorales, coquillages, poissons, selon un mode de vie côtier qui a pu favoriser une progression rapide vers l'est, jusqu'à l'Inde, l'Asie du Sud-Est et, finalement, le Sahul.
L'hypothèse de la route sud, dite aussi « dispersion côtière rapide », séduit par sa cohérence avec les données génétiques, qui suggèrent une expansion fulgurante le long des littoraux asiatiques. Mais elle reste difficile à documenter archéologiquement, car les sites côtiers de l'époque sont aujourd'hui ennoyés sous des dizaines de mètres d'eau, le niveau marin étant remonté depuis la dernière glaciation. Il est probable, en réalité, que les deux routes aient été empruntées à différentes époques, et que la sortie d'Afrique n'ait pas été un événement unique mais une série de pulsations migratoires conditionnées par les fluctuations du climat et de la végétation.
La preuve génétique : Ève mitochondriale et Adam du chromosome Y
Si la sortie d'Afrique a fini par s'imposer comme un paradigme, c'est avant tout grâce à la génétique des populations. Dès 1987, une équipe menée par Rebecca Cann, Mark Stoneking et Allan Wilson publia une étude fondatrice sur l'ADN mitochondrial. L'ADN mitochondrial, transmis exclusivement par la mère, ne se recombine pas : il se transmet de génération en génération en n'accumulant que des mutations ponctuelles, ce qui en fait une horloge moléculaire idéale pour remonter les lignées maternelles. En comparant l'ADN mitochondrial de populations du monde entier, les chercheurs constatèrent que toutes les lignées convergeaient vers un ancêtre commun unique, une femme ayant vécu en Afrique il y a quelque cent cinquante à deux cent mille ans : l'« Ève mitochondriale ».
Il importe de dissiper d'emblée un contresens tenace. L'Ève mitochondriale ne fut nullement la seule femme vivant à son époque, ni la « mère de l'humanité » au sens biblique. Elle est simplement la plus récente ancêtre commune par voie strictement maternelle de tous les humains actuels : des milliers d'autres femmes vivaient en même temps qu'elle, mais leurs lignées mitochondriales se sont éteintes au fil du temps, faute de filles à chaque génération. Un raisonnement symétrique, appliqué au chromosome Y transmis de père en fils, conduit à un « Adam du chromosome Y », lui aussi africain. Le fait majeur, pour notre propos, est que la diversité génétique humaine est maximale en Afrique et décroît à mesure que l'on s'en éloigne : c'est exactement ce que l'on attend si l'humanité s'est diffusée à partir d'un foyer africain, chaque vague de migrants n'emportant qu'un sous-échantillon de la variabilité de la population mère.
Cette signature, dite « sérielle » du goulot fondateur, constitue l'argument génétique le plus robuste en faveur de l'origine africaine récente. Plus une population est éloignée de l'Afrique au sens des routes migratoires, les Amérindiens, les Aborigènes d'Australie, moins elle est génétiquement diverse, car elle résulte d'une succession de fondations par de petits effectifs. La carte de la diversité génétique humaine est ainsi, littéralement, la carte de notre dispersion à partir du continent ancestral.
Le goulot d'étranglement et l'éruption du Toba
L'expansion d'Homo sapiens hors d'Afrique fut précédée et accompagnée d'un phénomène démographique majeur : un goulot d'étranglementGoulot d'étranglementRéduction brutale et temporaire de l'effectif d'une population, qui appauvrit durablement sa diversité génétique.→, c'est-à-dire une réduction drastique et temporaire de l'effectif de la population. Les analyses génétiques indiquent que les ancêtres de tous les non-Africains descendent d'un groupe fondateur étonnamment réduit, quelques milliers d'individus en âge de procréer, peut-être moins. Cet étranglement explique la relative pauvreté de la diversité génétique hors d'Afrique et la forte parenté de toutes les populations eurasiennes entre elles.
L'une des explications longtemps avancées pour ce goulot fut l'éruption du Toba. Il y a environ soixante-quatorze mille ans, le supervolcanSupervolcanVolcan capable de produire une éruption de magnitude 8 sur l'indice d'explosivité volcanique (IEV/VEI), c'est-à-dire émettant plus de 1 000 km³ de matériaux. Ces éruptions, dites « super-éruptions », sont extrêmement rares et laissent une caldeira géante plutôt qu'un cône.→ Toba, situé sur l'île de Sumatra, connut l'une des plus colossales éruptions des deux derniers millions d'années. Elle projeta des milliers de kilomètres cubes de matériaux, déposa une couche de cendres jusque sur le sous-continent indien et put provoquer un « hiver volcaniqueHiver volcaniqueRefroidissement global et prolongé du climat provoqué par l'injection d'aérosols sulfatés et de cendres dans la stratosphère lors d'une grande éruption, qui réfléchissent le rayonnement solaire et abaissent les températures pendant plusieurs années.→ » planétaire, un refroidissement brutal de plusieurs degrés ayant duré peut-être quelques années à quelques siècles. Selon la « théorie de la catastrophe de Toba », formulée par le géologue Stanley Ambrose, cet hiver aurait décimé les populations humaines, réduisant Homo sapiens au bord de l'extinction et créant précisément le goulot d'étranglement lu dans nos génomes.
Séduisante, cette hypothèse a néanmoins été fortement nuancée. Des fouilles menées en Inde et en Afrique australe, notamment sur des sites recouverts par les cendres du Toba, montrent une continuité des occupations humaines de part et d'autre de l'éruption, sans rupture nette. Par ailleurs, les estimations génétiques les plus récentes situent le goulot fondateur non pas tant au moment du Toba qu'au cœur même du processus de dispersion : le faible effectif des fondateurs serait avant tout la conséquence du petit nombre de migrants ayant réellement quitté l'Afrique et essaimé en Eurasie, indépendamment d'une quelconque catastrophe volcanique. Le Toba a sans doute affecté localement certaines populations, mais il n'est probablement pas le grand metteur en scène du goulot mondial qu'on a parfois voulu y voir.
Les métissages : Néandertal et Dénisoviens sur le chemin
Loin de migrer dans un monde vide, les sapiens sortis d'Afrique pénétrèrent un continent eurasien déjà peuplé par d'autres humanitésHominidéMembre de la lignée humaine au sens large, incluant les humains actuels, leurs ancêtres et les grands singes apparentés.→. En Europe et au Proche-OrientProche-OrientRégion d'Asie occidentale (Levant, Mésopotamie, Anatolie, Iran), berceau de la révolution néolithique, de l'agriculture, des premières villes et de l'écriture.→ vivaient les Néandertaliens, installés depuis des centaines de milliers d'années ; en Asie centrale et orientale rôdaient les DénisoviensDénisovienPopulation humaine éteinte, cousine des Néandertaliens, identifiée en 2010 par l'ADN de restes de la grotte de Denisova (Sibérie).→, cette population fantôme révélée par l'ADN d'une phalange sibérienne. Lorsque Homo sapiens rencontra ces lignées, il ne se contenta pas de les côtoyer ou de les remplacer : il s'unit à elles. L'hybridationHybridationCroisement entre deux espèces ou lignées distinctes, comme Homo sapiens et Néandertal, laissant une trace dans le génome.→ entre espèces humaines, longtemps tenue pour impossible ou tabou, est aujourd'hui un fait génétique établi.
Le séquençage du génome néandertalien, achevé en 2010 par l'équipe de Svante Pääbo, a révélé que tous les humains actuels non africains portent entre un et deux pour cent d'ADN néandertalien. Ce métissage s'est produit peu après la sortie d'Afrique, probablement au Proche-Orient, il y a environ cinquante à soixante mille ans, alors que les premiers migrants croisaient les Néandertaliens locaux. De même, les populations d'Océanie, Papous, Aborigènes australiens, Mélanésiens, portent jusqu'à quatre ou cinq pour cent d'ADN dénisovien, hérité de rencontres survenues en chemin, quelque part en Asie du Sud-Est, avec ces cousins encore mal connus.
Ces fragments archaïques ne sont pas de simples reliques inertes. Certains ont été conservés par la sélection naturelle parce qu'ils conféraient un avantage : des gènes néandertaliens liés à l'immunité, à la pigmentation ou au métabolisme, ou encore la fameuse variante du gène EPAS1, héritée des Dénisoviens, qui aide aujourd'hui les Tibétains à supporter le manque d'oxygène en haute altitude. La sortie d'Afrique ne fut donc pas seulement une diffusion, mais aussi une absorption : en se répandant, Homo sapiens a recueilli et intégré des fragments du patrimoine génétique des humanités qu'il rencontrait et qui, par ces traces, survivent encore en nous.
Le peuplement du monde : Europe, Asie, Sahul, Amériques
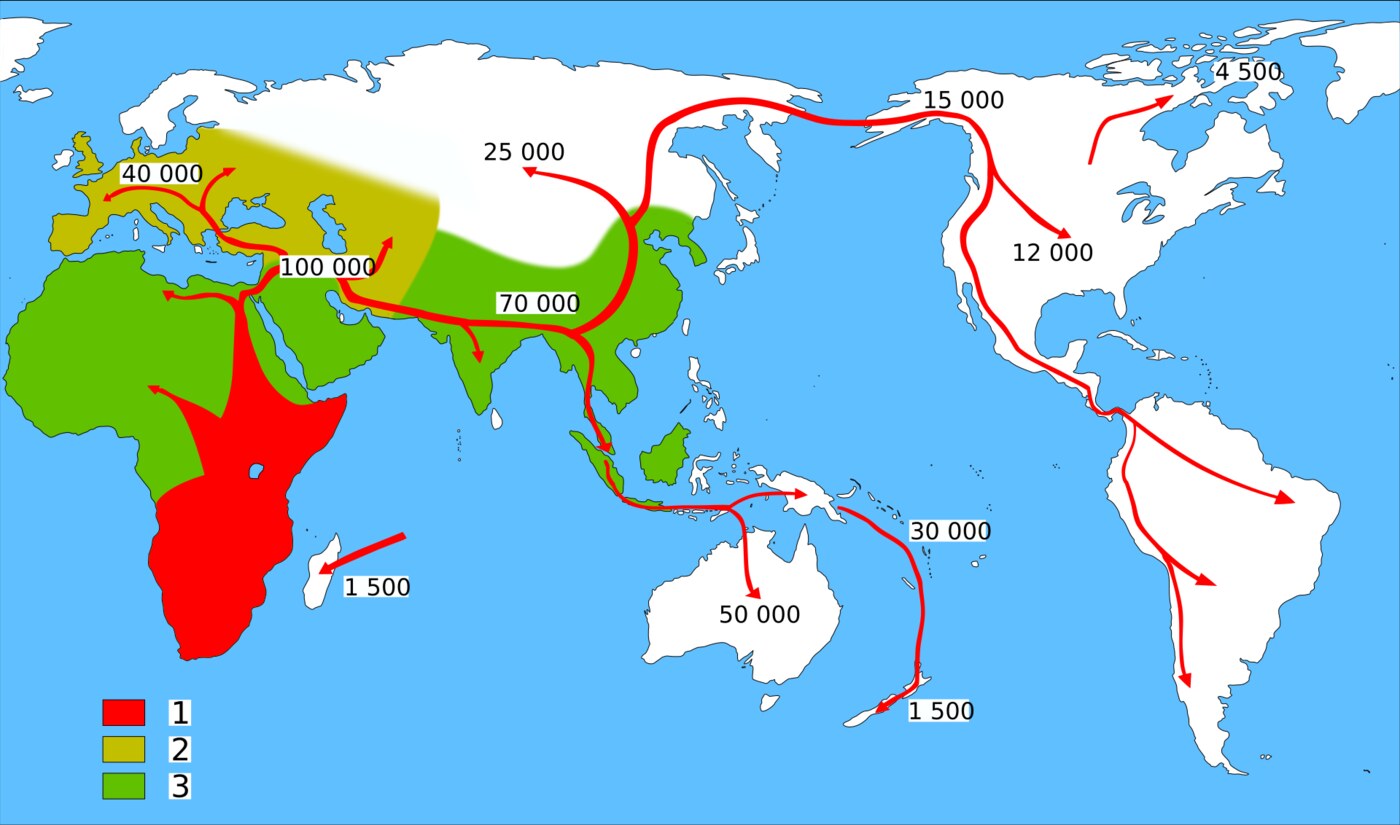
Une fois franchies les portes de l'Afrique, la dispersion d'Homo sapiens à travers le monde fut, à l'échelle du temps préhistorique, d'une rapidité saisissante. Vers quarante-cinq mille ans, les humains modernes atteignaient l'Europe, où ils sont associés à la culture aurignacienne et aux premières œuvres d'art figuratif, tout en cohabitant durant plusieurs millénaires avec les derniers Néandertaliens, avant l'extinction de ces derniers vers quarante mille ans. En Asie, la progression fut tout aussi vigoureuse, le long des côtes comme à l'intérieur des terres, jusqu'aux confins sibériens.
L'un des exploits les plus remarquables fut la colonisation du SahulSahulContinent formé, durant les glaciations, par la réunion de l'Australie, de la Nouvelle-Guinée et de la Tasmanie, lorsque le niveau marin était bas.→, ce continent qui réunissait, lors des bas niveaux marins du PléistocènePléistocèneÉpoque géologique des grandes glaciations (env. −2,6 Ma à −11 700), couvrant l'essentiel de la préhistoirePréhistoireEnsemble des périodes de l'histoire humaine antérieures à l'écriture, du Paléolithique à l'âge des métaux, connues principalement par les vestiges matériels.→ humaine.→, l'Australie, la Nouvelle-Guinée et la Tasmanie. Atteindre le Sahul depuis l'Asie du Sud-Est exigeait de franchir plusieurs bras de mer profonds, qui ne furent jamais entièrement asséchés même au plus fort des glaciations : c'est donc par la navigation, sur des embarcations dont nous ignorons tout, que les ancêtres des Aborigènes parvinrent en Australie, il y a au moins soixante-cinq mille ans selon les datations du site de Madjedbebe. Cette traversée maritime délibérée, la plus ancienne attestée de l'histoire humaine, témoigne de capacités cognitives et techniques pleinement modernes.
Le dernier grand continent à être peuplé fut l'Amérique. Par la BéringieBéringieVaste pont continental émergé entre la Sibérie et l'Alaska durant la dernière glaciation, à l'emplacement de l'actuel détroit de Béring ; steppe froide par laquelle les premiers Américains ont transité.→, ce pont terrestre qui reliait la Sibérie à l'Alaska lorsque le niveau marin était bas, des populations venues d'Asie pénétrèrent dans le Nouveau Monde, sans doute il y a entre vingt mille et seize mille ans, peut-être davantage selon des sites controversés comme White Sands, dont les empreintes pourraient remonter à plus de vingt mille ans. De l'Alaska à la Terre de Feu, ces pionniers descendirent l'ensemble des Amériques en quelques milliers d'années à peine. Ainsi, partie d'un foyer africain, l'humanité moderne avait, en l'espace de cinquante mille ans, atteint et habité la quasi-totalité des terres émergées de la planète.
Débats actuels
Le modèle de la sortie d'Afrique, pour solide qu'il soit dans ses grandes lignes, demeure un chantier scientifique en pleine effervescence. Plusieurs questions continuent de diviser les spécialistes. La première porte sur le nombre et la chronologie exacts des sorties : une dispersion unique tardive, ou plusieurs vagues échelonnées dont une seule aurait laissé une descendance ? Les fossiles de plus en plus anciens découverts en Asie et en Arabie compliquent le tableau et plaident pour un scénario à pulsations multiples.
La deuxième concerne l'origine africaine elle-même : le modèle panafricain de la métapopulation s'oppose encore à l'idée d'un berceau plus circonscrit, et la rareté des fossiles dans certaines régions du continent laisse de vastes zones d'ombre. La troisième touche au rôle exact du climat : on comprend de mieux en mieux que les « portes » de sortie s'ouvraient et se fermaient au rythme des cycles d'humidité et d'aridité, et la modélisation paléoclimatique devient un outil central pour reconstituer les fenêtres de migration possibles. Enfin, la question des métissages multiples s'enrichit sans cesse : on soupçonne désormais des introgressions plus anciennes et plus complexes, y compris des populations « fantômes » archaïques au sein même de l'Afrique, dont l'ADN aurait contribué au génome de certaines populations africaines actuelles.
Conclusion
La sortie d'Afrique n'est pas un simple épisode parmi d'autres de la préhistoire : c'est l'événement matriciel de l'unité humaine. Il nous rappelle que, par-delà l'extraordinaire diversité apparente des peuples, toutes les populations humaines actuelles partagent une origine commune et récente, et que les différences qui nous séparent sont infimes au regard de cet héritage partagé. Né en Afrique il y a trois cent mille ans, brassé à l'échelle d'un continent, sorti par les portes du Levant et de l'Arabie au gré des oscillations du climat, étranglé dans un goulot démographique puis enrichi par ses rencontres avec les Néandertaliens et les Dénisoviens, Homo sapiens a, en quelques dizaines de milliers d'années, conquis la planète entière.
Cette épopée, reconstituée pierre après pierre, gène après gène, demeure inachevée : chaque nouvelle fouille, chaque nouveau génome ancien séquencé en redessine les contours. Mais sa leçon centrale, elle, ne change pas. Nous sommes tous, sous toutes les latitudes, les descendants de ces voyageurs africains du PaléolithiquePaléolithiquePériode la plus ancienne et la plus longue de la préhistoire (env. −3,3 Ma à −12 000), définie par les outils de pierre taillée et un mode de vie de chasse et de cueillette.→ qui, sans en avoir conscience, suivant les troupeaux et les côtes, ont fait de la Terre entière le territoire d'une seule humanité.
Les origines africaines profondes
Avant de parler de sortie, il faut comprendre d'où l'on sort. L'émergence d'Homo sapiens ne fut pas un instant ponctuel mais un long processus enraciné dans l'épaisseur du temps africain. La découverte majeure de Jebel Irhoud, au Maroc, a bouleversé la chronologie admise : les restes humains exhumés sur ce site, longtemps datés de manière incertaine, ont été replacés par les méthodes de thermoluminescence à un âge moyen pondéré d'environ trois cent quinze mille ans, repoussant d'un coup de cent mille ans l'origine reconnue de notre espèce. Ces fossiles, associés à des outils du Middle Stone Age, présentent un visage déjà résolument moderne accolé à une boîte crânienne encore allongée, mosaïque de traits dérivés et de caractères plésiomorphesPlésiomorpheSe dit d'un caractère anatomique ancestral (primitif), hérité d'un ancêtre commun, par opposition aux traits dérivés récents.→ qui illustre à merveille la nature graduelle de notre devenir.
Jebel Irhoud ne forme cependant qu'une pièce d'un ensemble plus vaste. Le crâne de Florisbad, en Afrique du Sud, daté d'environ deux cent soixante mille ans, longtemps classé dans une zone grise taxonomique, a été rattaché à Homo sapiens archaïque à la lumière des nouvelles datations. Plus à l'est, en Éthiopie, les sites d'Omo Kibish et de Herto fournissent des jalons cruciaux : les restes d'Omo, autrefois estimés à cent quatre-vingt-quinze mille ans et récemment vieillis par la datation de couches volcaniques sus-jacentes, et ceux de Herto, datés d'environ cent soixante mille ans, comptent parmi les plus anciens fossiles incontestablement attribuables à notre espèce. Ensemble, ces gisements dispersés du Maghreb à la Corne de l'Afrique dessinent une géographie de l'émergence qui n'a rien d'un point unique sur la carte.
Cette dispersion même des plus anciens témoins pose une question de fond. Comment des populations présentant des traits modernes ont-elles pu apparaître à la fois au nord-ouest, au sud et à l'est du continent, séparées par des milliers de kilomètres et par des dizaines de milliers d'années ? La réponse, esquissée par la paléogénétiquePaléogénétiqueÉtude de l'ADN ancien extrait de restes (os, dents, sédiments, parois) pour reconstituer le passé des populations.→ et l'archéologie, oriente vers un modèle d'émergence diffuse plutôt que vers un foyer ponctuel, et c'est précisément ce modèle qui structure aujourd'hui le débat sur le berceau de l'humanité.
L'hypothèse panafricaine contre le berceau unique
Pendant des décennies, deux grands cadres se sont opposés pour expliquer notre origine. Le premier, le modèle de l'origine africaine récente dans sa version classique, postulait un berceau unique et géographiquement circonscrit, souvent placé en Afrique de l'Est, d'où une population fondatrice se serait répandue, remplaçant progressivement toutes les formes archaïques. Le second, dit panafricain ou multirégional africain, propose au contraire que Homo sapiens a émergé non pas en un lieu mais à l'échelle d'un continent tout entier, au sein d'une métapopulation fragmentée et reliée par des flux génétiques intermittents.
Selon cette vision panafricaine, des groupes d'Homo dispersés à travers les savanes, les forêts et les côtes africaines auraient échangé gènes et innovations culturelles au gré des connexions et des isolements imposés par les fluctuations climatiques. Tantôt réunis lors des phases humides qui verdissaient le Sahara, tantôt séparés par l'expansion des déserts, ces sous-ensembles auraient évolué en mosaïque, chacun contribuant pour partie au patrimoine de l'espèce en formation. Les traits modernes ne seraient alors pas apparus d'un seul tenant en un seul lieu, mais se seraient assemblés progressivement, brassés sur l'ensemble du continent, jusqu'à former le faisceau caractéristique de l'humanité actuelle.
Ce modèle reçoit aujourd'hui un appui considérable de la génétique des populations africaines, qui se révèlent porteuses d'une diversité bien supérieure à celle de toutes les autres populations du monde réunies. Cette profondeur généalogique trahit une histoire longue, structurée, faite de lignées anciennes qui ont divergé puis se sont parfois recroisées sur le sol africain. La distinction entre les deux modèles n'est d'ailleurs pas absolue : la plupart des chercheurs admettent désormais un compromis où l'Afrique de l'Est conserve un rôle de carrefour démographique majeur, sans pour autant être l'unique matrice d'où tout serait sorti. Le berceau de l'humanité serait moins un berceau qu'une vaste nurserie continentale.
Les sorties précoces : Misliya, Apidima, Skhul et Qafzeh
Si la grande dispersion qui peupla le monde se produisit relativement tard, elle fut précédée de plusieurs incursions plus anciennes, longtemps mésestimées et aujourd'hui reconnues comme des chapitres à part entière de l'histoire des sorties. Le plus spectaculaire de ces témoignages provient de la grotte de Misliya, sur le mont Carmel en Israël : une demi-mâchoire supérieure y a été datée d'une fourchette comprise entre cent soixante-dix-sept mille et cent quatre-vingt-quatorze mille ans, ce qui en fait le plus ancien fossile d'Homo sapiens connu hors d'Afrique et recule d'environ soixante mille ans la date du premier franchissement documenté du seuil africain.
D'autres indices renforcent ce tableau d'avant-garde. En Grèce, un fragment crânien de la grotte d'Apidima, daté d'environ deux cent dix mille ans, pourrait, selon certains chercheurs, appartenir à un sapiens très précoce, une attribution qui reste contestée, faute d'éléments diagnostiques suffisants, mais qui, si elle se confirmait, témoignerait d'une présence éphémère de notre espèce jusqu'au sud-est de l'Europe à une époque extraordinairement reculée. Plus tard, entre quatre-vingt-dix mille et cent vingt mille ans, les sépultures de Skhul et de Qafzeh, toujours au Levant, livrent des humains modernes incontestables, longtemps considérés comme les premiers émigrants africains avant la révélation de Misliya.
Ces incursions précoces partagent toutefois un trait décisif : elles semblent n'avoir laissé aucune descendance dans le patrimoine génétique des populations actuelles. Les hommes de Skhul, de Qafzeh ou de Misliya représentent vraisemblablement des avancées sans lendemain, des rameaux pionniers qui se sont éteints ou résorbés sans contribuer durablement au peuplement de l'Eurasie. Leur intérêt n'en est pas moindre : ils prouvent que la porte du Levant s'est ouverte et refermée à plusieurs reprises, au rythme des oscillations du climat, bien avant que la migration décisive ne s'engouffre dans cet interstice et n'essaime pour de bon.
La grande dispersion et ses preuves génétiques
Au-delà de ces tentatives avortées, c'est une migration plus tardive, située aux alentours de soixante mille ans, qui donna naissance à l'ensemble des populations non africaines actuelles. La convergence entre l'archéologie et la génétique est ici remarquable. L'analyse des lignées mitochondrialesADN ancienFragments d'ADN conservés dans des restes anciens (os, sédiments) ; leur séquençage permet d'identifier des espèces et de retracer des lignées disparues.→ montre que tous les non-Africains descendent de deux grands rameaux, les haplogroupes M et N, eux-mêmes issus d'un haplogroupe ancestral africain, L3, dont l'âge est estimé autour de soixante-dix mille ans. Cette concordance n'est pas fortuite : le même épisode démographique qui vit l'expansion de L3 en Afrique de l'Est paraît avoir alimenté la sortie d'un petit groupe fondateur qui allait peupler le reste du monde.
La jeunesse relative des lignées non africaines impose une conclusion forte : malgré les incursions anciennes attestées par les fossiles du Levant, l'écrasante majorité de l'ascendance des Eurasiens, des Océaniens et des Amérindiens remonte à cette unique dispersion réussie. La première expansion détectable génétiquement se situe entre cinquante-neuf mille et soixante-neuf mille ans, colonisant l'Asie occidentale et l'Inde avant d'atteindre rapidement l'Asie orientale. La faible diversité génétique observée chez les non-Africains, comparée à la richesse des populations restées en Afrique, signe la trace d'un goulot d'étranglementGoulot d'étranglementRéduction brutale et temporaire de l'effectif d'une population, qui appauvrit durablement sa diversité génétique.→ fondateur : seule une fraction réduite de la variabilité africaine a franchi le seuil et essaimé au-delà.
Cette signature génétique du goulot fondateur constitue l'un des piliers du modèle de la sortie d'Afrique. Elle explique pourquoi deux individus pris au hasard dans une population africaine peuvent différer davantage, dans leur ADN, que ne le font un Européen et un Asiatique de l'Est. Toute l'humanité extra-africaine descend d'une poignée de voyageurs, et cette origine étroite se lit encore, des dizaines de milliers d'années plus tard, dans chacun de nos génomes.
ADN mitochondrial et chromosome Y : Ève et Adam
Deux fragments singuliers du génome humain offrent une fenêtre privilégiée sur cette histoire des origines : l'ADN mitochondrial, transmis exclusivement par les mères, et le chromosome Y, transmis exclusivement de père en fils. Parce qu'ils échappent au brassage de la reproduction sexuée et se transmettent en bloc le long d'une seule lignée, ils permettent de remonter les généalogies maternelle et paternelle de l'humanité jusqu'à leurs ancêtres communs respectifs.
La généalogie maternelle converge vers une figure surnommée l'Ève mitochondriale, ancêtre commune matrilinéaire de tous les humains vivants, dont les estimations placent l'existence il y a environ cent cinquante mille à deux cent mille ans, certaines analyses récentes resserrant cette fourchette autour de cent à cent soixante mille ans. Du côté paternel, l'Adam du chromosome Y, ancêtre commun patrilinéaire, est situé selon les études entre cent vingt mille et deux cents mille ans. Il faut se garder d'un contresens tenace : ni l'Ève mitochondriale ni l'Adam chromosomique ne furent les seuls humains de leur temps, ni un couple primordial. Ils n'étaient que deux individus parmi une population nombreuse, simplement les seuls dont les lignées maternelle ou paternelle aient survécu sans interruption jusqu'à nous.
Ces deux ancêtres ont d'ailleurs vécu en Afrique, et probablement à des époques distinctes, séparées par des dizaines de milliers d'années, l'idée romantique d'un couple originel se dissout dans la statistique des coalescences génétiques. Leur intérêt est ailleurs : en datant ces points de convergence et en cartographiant la diversité des lignées qui en descendent, les généticiens reconstituent l'arbre généalogique de l'espèce, confirment l'enracinement africain de toutes les branches, et lisent dans la topologie de cet arbre la trace des grandes migrations. La plus grande profondeur des lignées africaines, tant maternelles que paternelles, y apparaît comme la marque indélébile du continent matriciel.
La route du nord et la route du sud
Par où, exactement, l'humanité sortit-elle d'Afrique ? Deux grands itinéraires ont été proposés, longtemps présentés comme rivaux et aujourd'hui considérés comme complémentaires. La route du nord empruntait le corridor levantin : remontant la vallée du Nil puis franchissant le Sinaï, les migrants débouchaient au Proche-Orient, ce carrefour où se sont retrouvés les fossiles de Misliya, de Skhul et de Qafzeh. C'est par cette voie que les premières incursions précoces semblent s'être faites, à la faveur des phases humides qui rendaient le désert du Sinaï et le Levant temporairement hospitaliers.
La route du sud, elle, franchissait le détroit de Bab-el-Mandeb, à l'extrémité méridionale de la mer Rouge, là où l'Afrique et la péninsule Arabique se font face à quelques dizaines de kilomètres seulement. Lors des bas niveaux marins du PléistocènePléistocèneÉpoque géologique des grandes glaciations (env. −2,6 Ma à −11 700), couvrant l'essentiel de la préhistoire humaine.→, ce passage se réduisait, et une traversée brève suffisait à gagner l'Arabie. Or l'image d'une Arabie éternellement désertique est trompeuse : à plusieurs reprises, le réchauffement des moussons a verdi la péninsule, multipliant lacs, rivières et savanes en une « Arabie verte » parsemée d'oasis. Ces fenêtres d'humidité ouvraient des couloirs de dispersion à travers ce qui n'est aujourd'hui qu'un océan de sable, et les découvertes archéologiques d'outils du Paléolithique dans le désert arabique attestent ces passages répétés.
Loin de s'exclure, les deux routes ont probablement fonctionné à des moments différents, chacune au gré des fenêtres climatiques qui s'entrouvraient. La porte du nord et la porte du sud battaient au rythme des cycles d'aridité et d'humidité, s'ouvrant lors des optima pour se refermer lors des phases sèches. La dispersion majeure aurait suivi un trajet largement côtier, longeant les rivages de l'océan Indien depuis l'Arabie jusqu'à l'Asie du Sud-Est, exploitant les ressources marines abondantes d'un littoral aujourd'hui largement submergé par la remontée des eaux post-glaciaires.
Le Toba et le débat du goulot d'étranglement
Au cœur de cette histoire se dresse l'ombre d'une catastrophe : l'éruption du supervolcan Toba, à Sumatra, survenue il y a environ soixante-quatorze mille ans, l'une des plus puissantes du Quaternaire. La théorie de la catastrophe de Toba proposait que cette super-éruption ait projeté dans l'atmosphère des volumes colossaux de cendres et d'aérosols, déclenchant un refroidissement global de plusieurs années et précipitant les effectifs humains sous le seuil de dix mille individus. Le goulot d'étranglement génétique observé chez l'humanité moderne aurait ainsi pour origine cette quasi-extinction volcanique.
Séduisante, cette hypothèse a néanmoins été fortement nuancée, voire contestée. Des fouilles menées en Inde et en Afrique australe, notamment sur des sites recouverts par les cendres du Toba, montrent une continuité des occupations humaines de part et d'autre de l'éruption, sans rupture nette du peuplement. Des populations ont survécu et sont demeurées actives après la catastrophe, ce qui affaiblit l'idée que cet événement aurait été la cause principale du goulot fondateur. Par ailleurs, les estimations génétiques les plus récentes situent ce goulot non pas tant au moment du Toba qu'au cœur même du processus de dispersion : le faible effectif des fondateurs serait avant tout la conséquence du petit nombre de migrants ayant réellement quitté l'Afrique, indépendamment de toute catastrophe volcanique.
Le débat conserve toutefois son acuité. Que la date du Toba précède de peu l'expansion de l'haplogroupe L3 n'est sans doute pas un pur hasard, et certains chercheurs maintiennent qu'un refroidissement, même bref, a pu remodeler les populations et reconfigurer les fenêtres de migration. Le Toba a certainement affecté localement certaines régions ; mais il n'est probablement pas le grand metteur en scène du goulot mondial qu'on a parfois voulu y voir. Le resserrement démographique des fondateurs paraît avant tout inhérent à la mécanique même de la dispersion, qu'aucune catastrophe ne fut nécessaire pour produire.
Les adaptations à de nouveaux mondes
En se répandant sous toutes les latitudes, Homo sapiens rencontra des environnements radicalement étrangers à ceux de la savane tropicale qui l'avait vu naître : déserts froids, hauts plateaux, forêts boréales, régions peu ensoleillées. La sélection naturelle, opérant à des rythmes accélérés sur ces fronts pionniers, façonna en quelques milliers de générations un éventail d'adaptations locales qui se lisent encore dans la diversité physiologique actuelle. La pigmentation cutanée en offre l'exemple le plus visible : sous les latitudes peu ensoleillées d'Eurasie, des variantes des gènes de pigmentation, tels SLC24A5 et SLC45A2, furent sélectionnées pour éclaircir la peau et favoriser la synthèse de vitamine D, tandis que la peau sombre des origines protégeait des ultraviolets intenses des tropiques.
D'autres adaptations témoignent de la plasticité de l'espèce face à des défis spécifiques. La persistance de la lactase, qui permet de digérer le lactose à l'âge adulte, est apparue de façon indépendante dans plusieurs populations d'éleveurs, en Europe comme en Afrique, par des mutations distinctes du gène LCT, un cas d'école d'évolution convergente liée au pastoralismePastoralismeMode de vie fondé sur l'élevage de troupeaux (bovins, ovins, caprins), souvent mobile, qui s'est répandu au Sahara « vert » et a précédé, dans cette région, l'agriculture proprement dite.→. En haute altitude, sur le plateau tibétain où l'air se raréfie au-dessus de quatre mille mètres, la variante du gène EPAS1, héritée des DénisoviensDénisovienPopulation humaine éteinte, cousine des Néandertaliens, identifiée en 2010 par l'ADN de restes de la grotte de Denisova (Sibérie).→ lors d'une hybridationHybridationCroisement entre deux espèces ou lignées distinctes, comme Homo sapiens et Néandertal, laissant une trace dans le génome.→ ancienne, module la production de globules rouges et permet de tolérer l'hypoxie sans les effets délétères d'une surproduction sanguine.
Le métabolisme lui-même porte la marque de ces ajustements : des variantes des gènes FADS, impliqués dans la transformation des acides gras, furent sélectionnées en fonction des régimes alimentaires locaux, qu'ils fussent riches en ressources marines, terrestres ou végétales. Ces adaptations, dispersées dans le génome, ne séparent en rien l'humanité en groupes étanches : elles forment une marqueterie de réponses ponctuelles à des pressions environnementales précises, surimposées à un fonds génétique commun écrasant. La diversité visible des peuples n'est que la fine pellicule adaptative d'une unité profonde.
Ce que révèle la génomique des populations actuelles
La paléogénétique ne se nourrit pas seulement d'os fossiles ; elle interroge aussi, et avec une fécondité croissante, le génome des populations vivantes. Le séquençage à grande échelle de milliers d'individus issus de tous les continents a confirmé et raffiné le tableau de la sortie d'Afrique. Le constat le plus robuste est celui d'un gradient de diversité décroissant à mesure que l'on s'éloigne de l'Afrique : plus une population est géographiquement distante du continent matriciel, plus sa diversité génétique est appauvrie, conformément au modèle des goulots successifs qui rabotaient la variabilité à chaque nouvelle étape de la colonisation.
Cette structure en éventail, enracinée en Afrique et s'amincissant vers les confins du peuplement, constitue la signature même d'une expansion à partir d'un foyer unique. Le génome des populations actuelles enregistre par ailleurs, comme un palimpseste, les traces des métissages anciens : le pourcentage d'ADN néandertalien chez les Eurasiens, la fraction dénisovienne chez les Océaniens, et jusqu'à des introgressions plus obscures, attribuées à des populations « fantômes » dont nous ne connaissons aucun fossile mais dont l'ADN subsiste dans certaines lignées africaines actuelles.
La génomique des populations vivantes éclaire enfin les routes de la dispersion. La distribution géographique des haplogroupes, la datation de leurs ramifications, la cartographie des affinités entre groupes humains permettent de reconstituer les itinéraires, les vitesses et les directions de la grande migration. Chaque génome individuel est ainsi une archive : il porte, inscrite dans la séquence de ses bases, la mémoire des voyages de ses ancêtres, depuis le berceau africain jusqu'aux extrémités habitées de la planète. La biologie moléculaire est devenue, à sa manière, une science historique.
Incertitudes et perspectives
Pour solide qu'il soit dans ses grandes lignes, le récit de la sortie d'Afrique demeure un chantier ouvert, traversé de zones d'ombre et de controverses fécondes. La chronologie fine des départs continue de se complexifier à mesure que de nouveaux fossiles, toujours plus anciens, sont mis au jour en Asie et en Arabie : faut-il imaginer une seule dispersion tardive et décisive, ou une succession de pulsations migratoires dont une seule aurait essaimé durablement ? Les données génétiques penchent pour la seconde hypothèse, sans exclure que des traces ténues de migrations plus anciennes subsistent dans certains génomes asiatiques, à l'état de murmures noyés dans le flux dominant.
Nous sommes tous, sous toutes les latitudes, les héritiers d'une même odyssée africaine ; la diversité des peuples n'est que la surface d'une parenté abyssale.
D'autres incertitudes pèsent sur le tableau. Le rôle exact du climat dans l'ouverture et la fermeture des « portes » de sortie se précise grâce à la modélisation paléoclimatique, devenue un instrument central pour identifier les fenêtres de migration possibles, mais la résolution de ces modèles reste grossière au regard de la finesse des événements démographiques. La question des métissages, elle, ne cesse de s'approfondir : on soupçonne des introgressions plus nombreuses, plus anciennes et plus complexes que ne le laissait croire le schéma initial, y compris au sein même de l'Afrique. Chaque génome ancien séquencé, chaque couche de sédiment analysée pour son ADN environnementalADN environnementalADN libéré par les organismes dans leur milieu (sol, sédiment, eau, paroi) et récupérable sans le moindre reste corporel identifiable.→, redessine un peu les contours de l'épopée. Loin d'ébranler la trame générale, ces incertitudes en confirment la vitalité : la sortie d'Afrique n'est pas un dogme figé mais une science en marche, dont les prochaines découvertes affineront encore le récit sans en renverser la leçon centrale.
Au terme de ce parcours, une certitude domine toutes les controverses de détail. Quelles qu'aient été les routes empruntées, le nombre exact des vagues ou la part respective des catastrophes et du hasard démographique, l'humanité contemporaine forme un seul peuple dispersé. Née en Afrique au sein d'une métapopulation continentale il y a quelque trois cent mille ans, sortie par les seuils du Levant et de Bab-el-Mandeb au gré des oscillations du climat, étranglée dans un goulot fondateur puis enrichie par ses rencontres avec d'autres humanitésHominidéMembre de la lignée humaine au sens large, incluant les humains actuels, leurs ancêtres et les grands singes apparentés.→, notre espèce a tissé en quelques dizaines de millénaires une toile de peuplement couvrant la quasi-totalité des terres émergées. La paléogénétique et l'archéologie, en confrontant sans cesse leurs résultats, achèvent de démontrer ce que les anciens récits pressentaient à leur manière : par-delà l'apparente mosaïque des visages et des langues, c'est une seule et même lignée de voyageurs du PaléolithiquePaléolithiquePériode la plus ancienne et la plus longue de la préhistoire (env. −3,3 Ma à −12 000), définie par les outils de pierre taillée et un mode de vie de chasse et de cueillette.→ qui se reconnaît, aujourd'hui, dans le miroir de son propre génome.
La théorie Out of Africa reste le cadre interprétatif dominant mais elle doit aujourd'hui intégrer des données de plus en plus complexes. La découverte de fossiles d'Homo sapiens hors d'Afrique datés de plus de 100 000 ans à Misliya en Israel et dans la péninsule arabique suggère des sorties précoces qui ne correspondent pas toujours à la colonisation finale du globe. Le débat est loin d'être clos.
Le modèle Out of Africa a été largement confirmé par la génétique des populations mais il se révèle beaucoup plus complexe qu'initialement proposé. Les données génomiques modernes et anciennes suggèrent des sorties d'Afrique multiples, des retours possibles vers l'Afrique, et des métissages avec des populations archaïques en Asie et en Europe. Le modèle de remplacement complet est remplacé par un modèle d'assimilation partielle.